
Two or three types of cone photoreceptor and a single type of rod photoreceptor are present in the normal mammalian retina. Some non-mammalian retinas have even more cone types (see later).
1. Light microscopy and ultrastructure of rods and cones
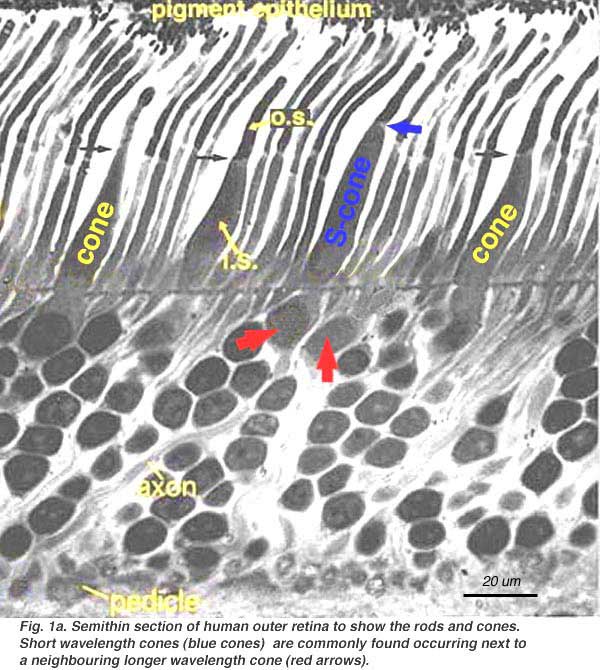
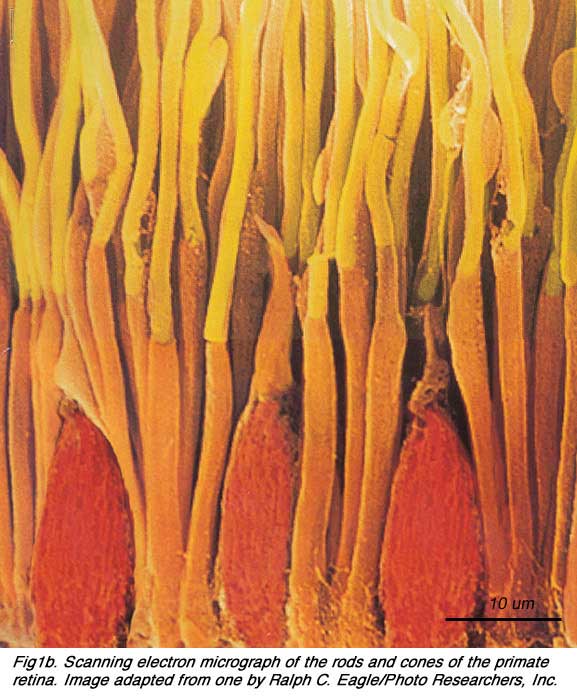
In vertical sections of retina prepared for light microscopy with the rods and cones nicely aligned, the rods and cones can be distinguished rather easily.
 |  |
{kind=link}
{kind=link}
Cones are robust conical-shaped structures that have their cell bodies situated in a single row right below the outer limiting membrane (OLM) and their inner and outer segments protruding into the subretinal space towards the pigment epithelium (Figs. 1 and 2). In foveal retina, where only cones are concentrated, their cell bodies are layered in oblique columns below the outer limiting membrane. Rods, on the other hand, are slim rod-shaped structures with their inner and outer segments filling the area between the larger cones in the subretinal space and stretching to the pigment epithelium cells. Rod cell bodies make up the remainder of the outer nuclear layer below the cone cell bodies. Apical processes from the pigment epithelium envelope the outer segments of both rods and cones (not always clear in histological sections).
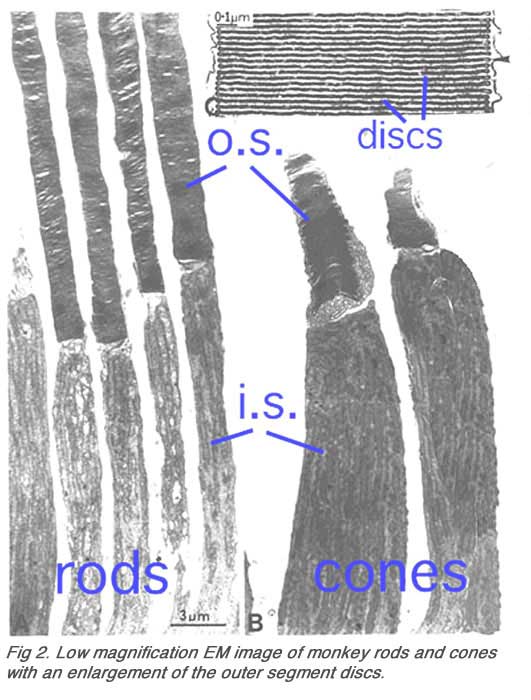
The higher magnification afforded by the electron microscope allows better resolution of rod and cone photoreceptors.
 |  |
{kind=link}
{kind=link}
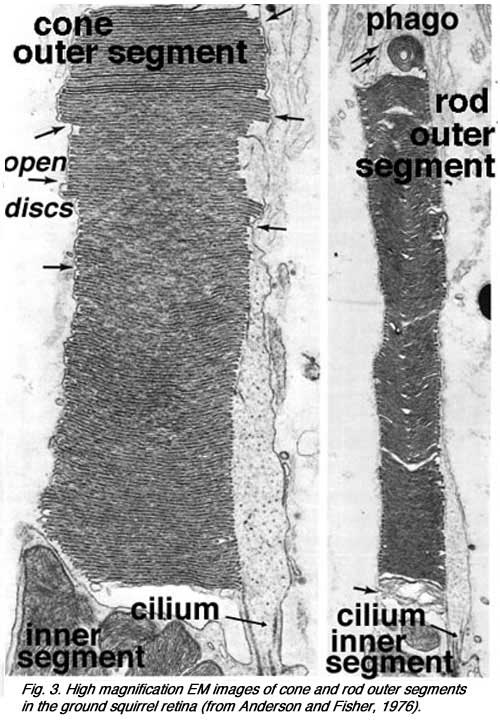
Ultrathin sections viewed in an electron microscope (Figs. 2 and 3) show rods and cones from human and squirrel retinas (Anderson and Fisher, 1976). The photoreceptor consists of 1) an outer segment, filled with stacks of membranes (like a stack of poker chips) containing the visual pigment molecules such as rhodopsins, 2) an inner segment containing mitochondria, ribosomes and membranes where opsin molecules are assembled and passed to be part of the outer segment discs, 3) a cell body containing the nucleus of the photoreceptor cell and 4) a synaptic terminal where neurotransmission to second order neurons occurs.
CLICK to see a photoreceptor with emphasis on the inner and outer segments and the disc structures within the outer segment. Movie made by Carlos Rozas (CanalWeb, Chile). 2 MB Quicktime movie.
Outer and inner segments of rods (i.s.) are generally thinner than those of cones in mammalian retinas. For example, the rod inner segments are 2 microns and the cone’s about 6 microns in diameter in peripheral human retina. In the fovea, however, where there are only cone photoreceptors, the most central cones are even thinner than the average rod at about 1.5 microns diameter. Inner segment regions of both rods and cones are filled with long thin mitochondria. At the top of the inner segment a thin cilium joins the inner and outer segments of the rods and cones (Fig. 3 and see movie).
2. Outer segment generation
It is from the base of the cilium that membrane evaginations and invaginations occur to produce the outer segment (o.s.) or the important visual pigment-bearing portion of the photoreceptor. Outer segments of both the rods and cones arises from outpouching (a, Fig. 5 below) of the photoreceptor cell plasma membrane at this point (see below) (Steinberg et al., 1980).
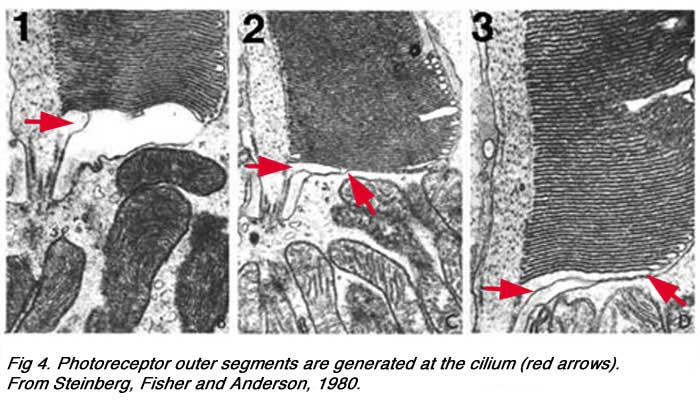 | 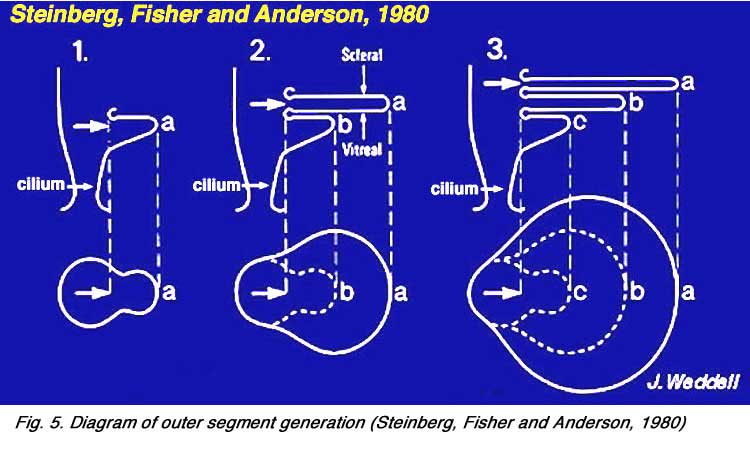 |
{kind=link}
{kind=link}
CLICK to see an animation of the outer segment generation
(420 K quicktime movie)
These expanding membrane plates (b-c, Fig. 5 above) become detached as free floating discs inside the outer segment membrane in the case of the rods. In the case of the cones though, the outer segment discs remain attached to the outer segment membrane.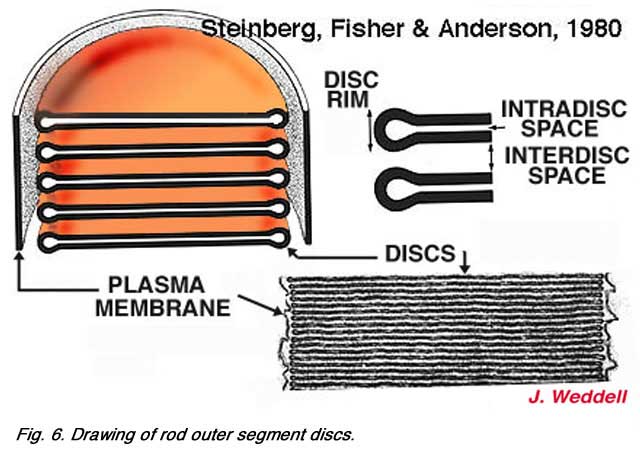 So the outer segment is a structure filled entirely with discs of folded double membranes in which are embedded the light sensitive visual pigment molecules (Fig. 6).
So the outer segment is a structure filled entirely with discs of folded double membranes in which are embedded the light sensitive visual pigment molecules (Fig. 6).
The opsin molecule, which binds the chromophore 11-cis retinal to form the visual pigment, is manufactured in the Golgi apparatus of the inner segment and presented to the outer membrane at the cilium, via fusion areas using G-proteins (Papermaster et al., 1985; Deretic and Papermaster, 1995).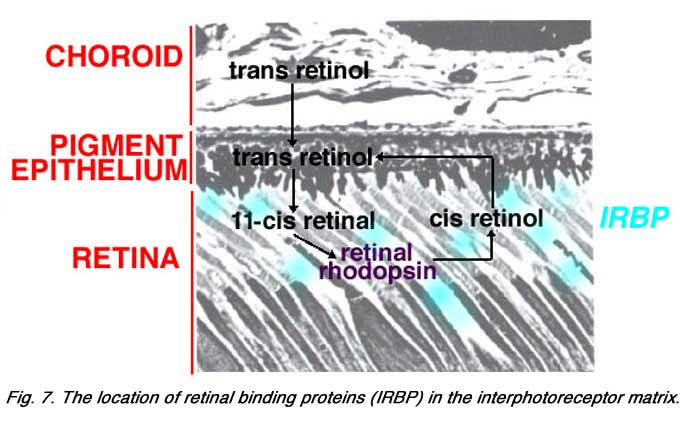 In contrast, the other part of the visual pigment molecule in the outer segment discs which is retinal (vitamin A product), is provided to the discs from the pigment epithelium via carrier molecules (retinal binding proteins, IRBP) in the interphotoreceptor matrix of the subretinal space (Fig. 7) (Adler and Martin, 1982; Chader, 1989) and see chapter on retinal pigment epithelium by Olaf Strauss.
In contrast, the other part of the visual pigment molecule in the outer segment discs which is retinal (vitamin A product), is provided to the discs from the pigment epithelium via carrier molecules (retinal binding proteins, IRBP) in the interphotoreceptor matrix of the subretinal space (Fig. 7) (Adler and Martin, 1982; Chader, 1989) and see chapter on retinal pigment epithelium by Olaf Strauss.
3. Visual pigments and visual transduction
Vertebrate photoreceptors can respond to light by virtue of their containing a visual pigment embedded in the bilipid membranous discs that make up the outer segment. The visual pigment consists of a protein called opsin and a chromophore derived from vitamin A known as retinal. The vitamin A is manufactured from beta-carotene in the food we eat, and the protein is manufactured in the photoreceptor cell (see above). The opsin and the chromophore are bound together and lie buried in the membranes of the outer segment discs (Fig. 8).
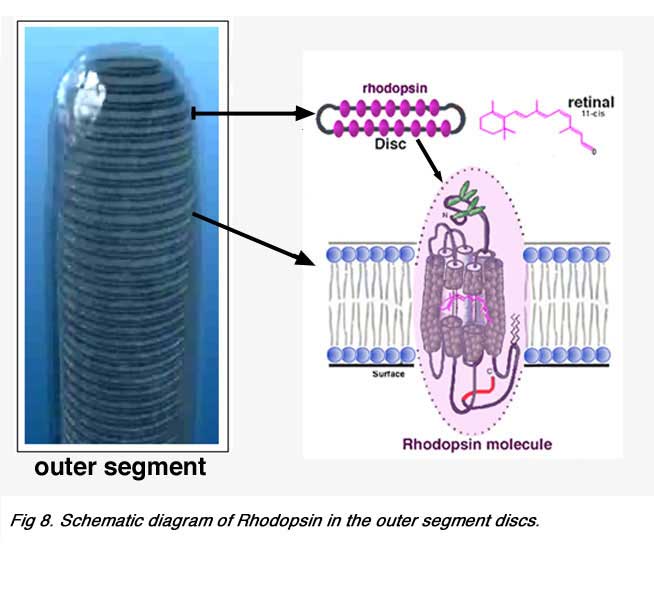 |  |
{kind=link}
{kind=link}
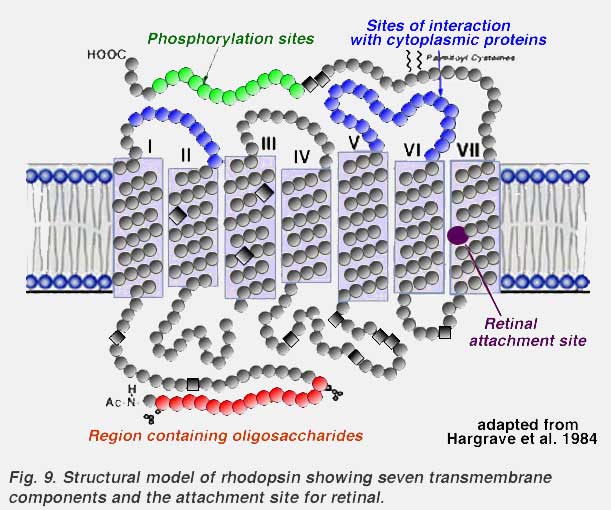
About 50% of the opsin is within the bilipid membrane connected by short protein loops outside. Each molecule of rhodopsin consists of seven of these transmembrane portions surrounding the chromophore (11-cis retinal) in the lipid bilayer (Fig. 9). The chromophore apparently lies horizontally in the membrane and is bound at a lysine residue to helix seven (Hargrave et al. 1984, Hargrave and McDowell, 1992)(Fig. 9). Each outer segment disc, of course, contains many (thousands) visual pigment molecules. Upon absorption of a photon of light, the retinal isomerizes from the 11-cis form to an all-trans form which starts conformational changes in the molecule resulting in bleaching. Several intermediaries are formed in bleaching among them metarhodopsin II which activates the G-protein transducin and a further cascade of events summarized below (see review by Hargrave and McDowell (1992) and by Archer, 1995) and chapter by Yingbin Fu (webvision).
Light transduces the visual pigment via the following enzyme cascade: photons – rhodopsin – activated rhodopsin (metarhodopsin II) – a GTP binding protein (transducin) – an enzyme hydrolyzing cGMP (cGMP-phosphodiesterase) – closes a membrane bound cGMP-gated cation channel.
In the dark a steady current flows into the open channels, carried mainly by Na ions, constituting a “dark current” that partially depolarizes the photoreceptor cell (Fig. 10). Thus, the depolarized photoreceptor releases neurotransmitter (the amino acid glutamate) from its synaptic terminals upon second-order neurons in the dark. On light stimulation the rhodopsin molecules are isomerized to the active form, the above cascade ensues, leading to closure of the cation channels of the photoreceptor membrane, stopping the dark current and causing the photoreceptor cell membrane to hyperpolarize and cease neurotransmitter release to second-order neurons (Fig. 10) (see Stryer, 1991; Yau, 1994, and Kawamura, 1995, and Fu (webvision) for reviews).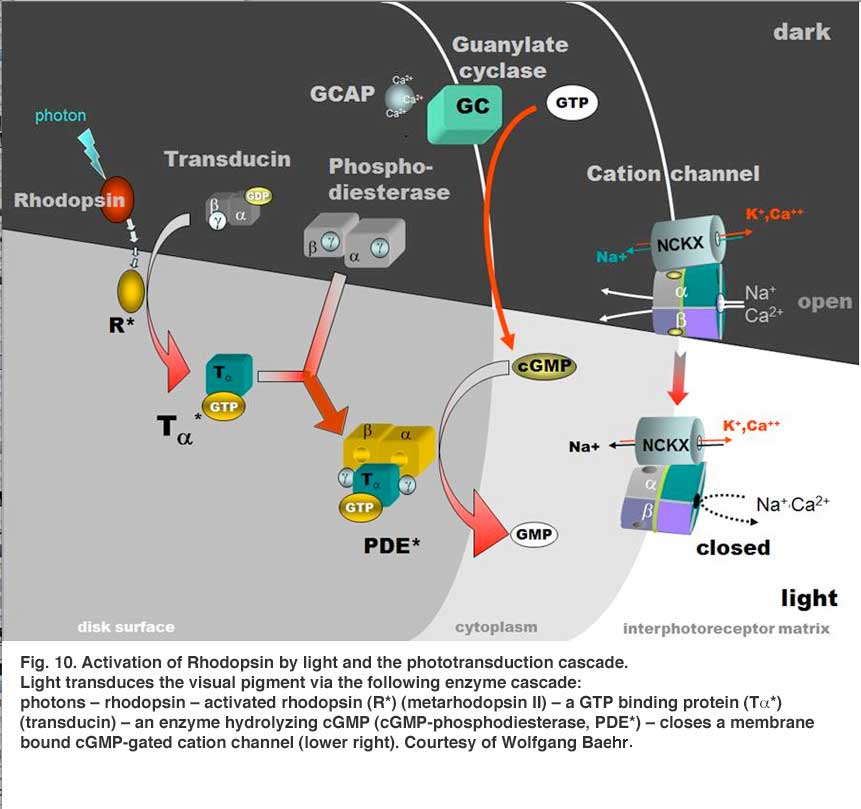
Click to see an animation of phototransduction (800K quicktime movie)
The “dark current” is composed mainly of the influx of the Na+ component (80%) however, a Ca2+ component (15%) and a Mg2+ component (5%) are also present (Yau, 1994). In darkness there must be a mechanism to remove Ca2+ as well as the excess Na+, and it is thought to be done so through a sodium/calcium exchanger in the membranes of the outer segment of the photoreceptor. Ca2+, once thought to be the second messenger in linking the rhodopsin photoisomerization to the membrane events is now known to have a secondary but important regulatory role in phototransduction. Although it does not directly participate in the transduction cascade it does improve the signalling capability of the rods in speeding the recovery after illumination and down regulating of the rods sensitivity in steady illumination (Yau, 1994). The latter effect is a mechanism for light adaptation.
For it must be remembered that a photoreceptor cell does not simply detect light. It can also adapt to environmental light. For example cone photoreceptors can adapt so that our visual system can see from the dim shadows under a tree to objects in bright sunlight snow, a shift of light intensity of 7-9 log units of light intensity (Normann et al., 1991). Rod photoreceptors, thought at one time not to light adapt, are now known to adapt over a range of 2 log units of background intensity and combined with a network adaptation through the whole visual system, allows as much as 5 log units of background intensity adaptation in rod driven vision (Yau, 1994).
4. Phagocytosis of outer segments by pigment epithelium
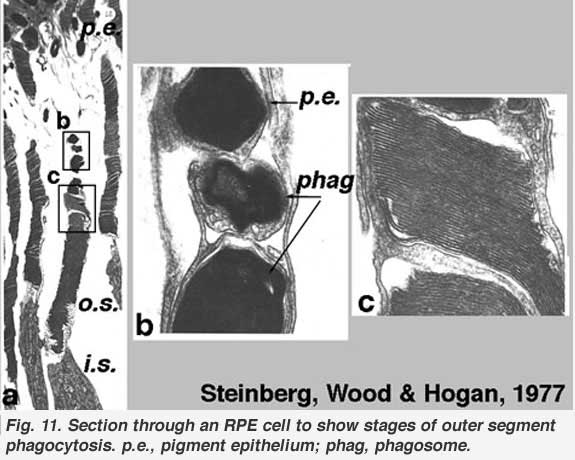
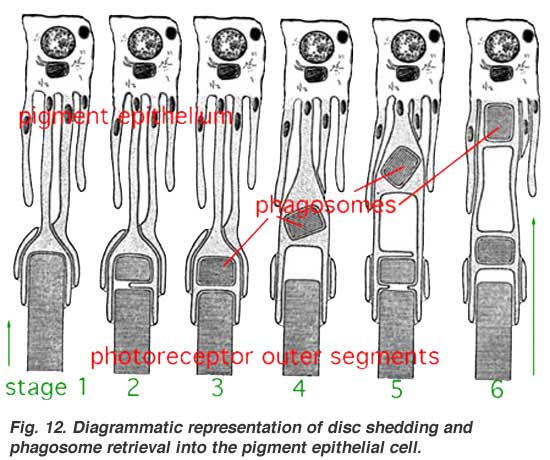
The stacks of discs containing visual pigment molecules in the outer segments of the photoreceptors are constantly renewed. New discs are added at the base of the outer segment at the cilium as discussed above. At the same time old discs are displaced up the outer segment and are pinched off at the tips and engulfed by the apical processes of the pigment epithelium (Figs. 11 and 12). These discarded, spent discs become known as phagosomes in the pigment epithelial cells. They are then broken down by lysis. Photoreceptor outer segment discs are phagocytosed by the pigment epithelium in a diurnal cycle. There is a burst of disc shedding at light on in the morning, judged by increased numbers of phagosomes in the pigment epithelium shortly thereafter (Young 1971).
Click to see an animation of the phagocytosis (956 K quicktime movie)
 |  |
{kind=link}
{kind=link}
Cone outer segments differ from rod outer segments in several respects. Firstly, they are shorter and more conical with a wider base and tapering shape compared with those of rods. Secondly, as mentioned above, their discs are connected to the plasma membrane throughout the extent of the outer segment, and thus they are open to extracellular space. Apical processes of the pigment epithelium phagocytize chunks of cone outer segments, just like they do the rod outer segments, but at a different time in the diurnal cycle compared with rods i.e. at light off-set compared light-onset (see original work by Young, 1971, 1976; LaVail, 1976; Steinberg et al., 1977; reviewed by Besharse, 1982).
5. Different types of cone photoreceptor
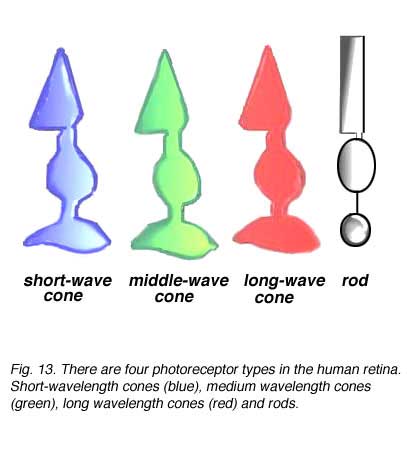 As we have seen from the morphological appearances described above, two basic types of photoreceptor, rods and cones, exist in the vertebrate retina (Fig. 13). The rods are photoreceptors that contain the visual pigment – rhodopsin and are sensitive to blue-green light with a peak sensitivity around 500 nanometers (nm) wavelength of light (Fig. 14a). Rods are highly sensitive photoreceptors and are used for vision under dark-dim conditions at night. Cones contain cone opsins as their visual pigments and, depending on the exact structure of the opsin molecule, are maximally sensitive to either long wavelengths of light (red light), medium wavelengths of light (green light) or short wavelengths of light (blue light). Cones of different wavelength sensitivity and the consequent pathways of connectivity to the brain are, of course, the basis of color perception in our visual image.
As we have seen from the morphological appearances described above, two basic types of photoreceptor, rods and cones, exist in the vertebrate retina (Fig. 13). The rods are photoreceptors that contain the visual pigment – rhodopsin and are sensitive to blue-green light with a peak sensitivity around 500 nanometers (nm) wavelength of light (Fig. 14a). Rods are highly sensitive photoreceptors and are used for vision under dark-dim conditions at night. Cones contain cone opsins as their visual pigments and, depending on the exact structure of the opsin molecule, are maximally sensitive to either long wavelengths of light (red light), medium wavelengths of light (green light) or short wavelengths of light (blue light). Cones of different wavelength sensitivity and the consequent pathways of connectivity to the brain are, of course, the basis of color perception in our visual image.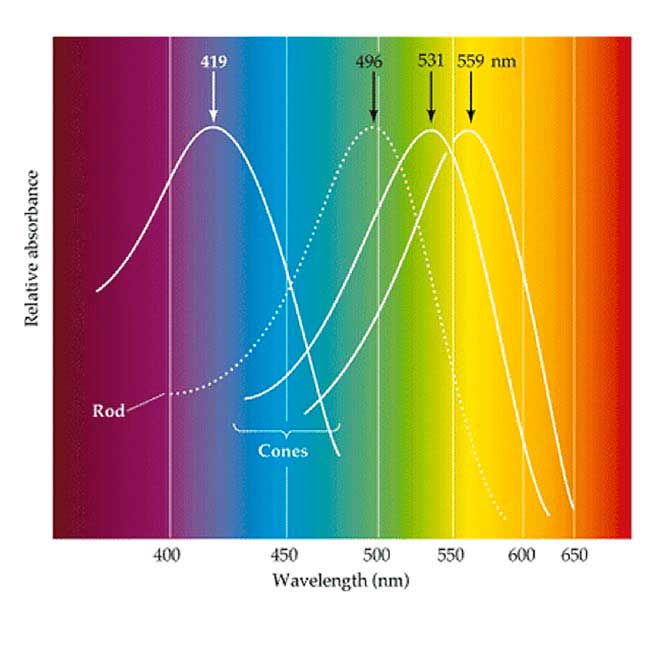 Three different cone mechanisms can be detected in behavioral, psychophysical, physiological and molecular staining in primate retinas (Fig. 14a). This 3-cone mechanism is the basis of so called trichromatic vision which most humans have. Thus, long wavelength sensitive, L-cones (red) are known to be maximally sensitive to wavelengths peaking around 560nm, medium wavelength sensitive, M-cones (green) peak around 530nm and short wavelength cones, S-cones (blue) peak at 420nm respectively (Fig. 14a) (Bowmaker and Dartnell, 1980; Bowmaker et al., 1980, and see chapter on color vision in webvision). Other authors have a different result for the peak sensitivity of the blue cones, or S-cones, with peaks varying from 430nm (Baylor et al., 1987) to 445nm (Dobelle, Marks and MacNichol 1969).
Three different cone mechanisms can be detected in behavioral, psychophysical, physiological and molecular staining in primate retinas (Fig. 14a). This 3-cone mechanism is the basis of so called trichromatic vision which most humans have. Thus, long wavelength sensitive, L-cones (red) are known to be maximally sensitive to wavelengths peaking around 560nm, medium wavelength sensitive, M-cones (green) peak around 530nm and short wavelength cones, S-cones (blue) peak at 420nm respectively (Fig. 14a) (Bowmaker and Dartnell, 1980; Bowmaker et al., 1980, and see chapter on color vision in webvision). Other authors have a different result for the peak sensitivity of the blue cones, or S-cones, with peaks varying from 430nm (Baylor et al., 1987) to 445nm (Dobelle, Marks and MacNichol 1969).
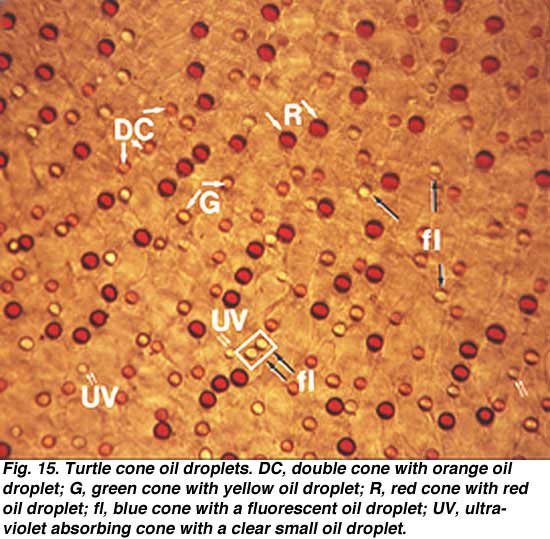
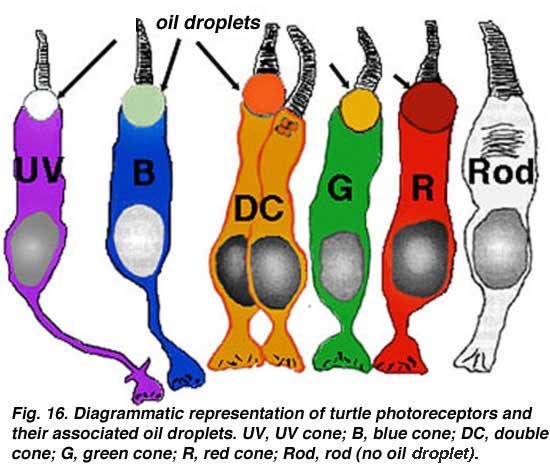
Normal human color vision depends on these three cone mechanisms. This adds an added dimension to color vision over that of mammals which are mostly dichromatic. Dichromats have just long wavelength (red) and short wavelength (blue) cones. In primates and humans, nature has split the long-wave system into two systems with slightly different spectral sensitivities. The blue cones are like those found in dichromatic mammals. Fig. 14b shows the subtle difference in the molecular structure of the red, green and blue cone photopigments compared with the rhodopsin rod pigment (different molecules are colored pink (Nathans et al., 1986).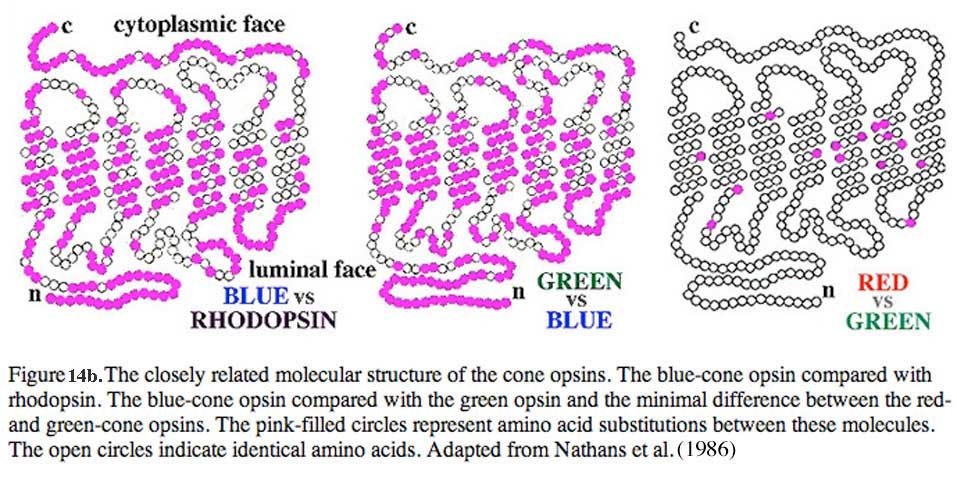 To understand color vision and how the colored visual message is processed in the retina we need to be able to start with a morphological distinction of the three (or more) cone types, so they can then, hopefully be identified with any color specific connections they make, i.e. connections to bipolar, horizontal cell and finally ganglion cells of the retina. Fortunately, certain vertebrate species have distinctly different morphological cone types in their retinas and it has recently become possible to correlate these morphologies with spectral sensitivity. Now we can distinguish short, medium and long wavelength cones in retinas of some fish, frogs, birds, and reptiles (turtles) based on distinct morphological differences. Turtle retinas for example have colored oil droplets in their different spectral types of cone which identify them rather readily (Figs. 15 and 16) (see reviews by Kolb and Lipetz, 1991 and Ammermüller and Kolb, 1996).
To understand color vision and how the colored visual message is processed in the retina we need to be able to start with a morphological distinction of the three (or more) cone types, so they can then, hopefully be identified with any color specific connections they make, i.e. connections to bipolar, horizontal cell and finally ganglion cells of the retina. Fortunately, certain vertebrate species have distinctly different morphological cone types in their retinas and it has recently become possible to correlate these morphologies with spectral sensitivity. Now we can distinguish short, medium and long wavelength cones in retinas of some fish, frogs, birds, and reptiles (turtles) based on distinct morphological differences. Turtle retinas for example have colored oil droplets in their different spectral types of cone which identify them rather readily (Figs. 15 and 16) (see reviews by Kolb and Lipetz, 1991 and Ammermüller and Kolb, 1996).
 |  |
{kind=link}
{kind=link}
However, primate and human retinas still contain cone types which look essentially the same morphologically, but here too with the latest anatomical techniques we are beginning to be able to see at least a difference between the short wavelength cone and the two longer wavelength cones. Specialized histochemical techniques (Marc and Sperling, 1977), dye uptake studies (DeMonasterio et al., 1981) or use of antibodies specific for visual pigments (Szel et al., 1988) have allowed identification of the different spectral types of cone in most mammalian species now. In primate retina antibodies against visual pigments stain outer segments of the L/M-cones together or the S-cones only.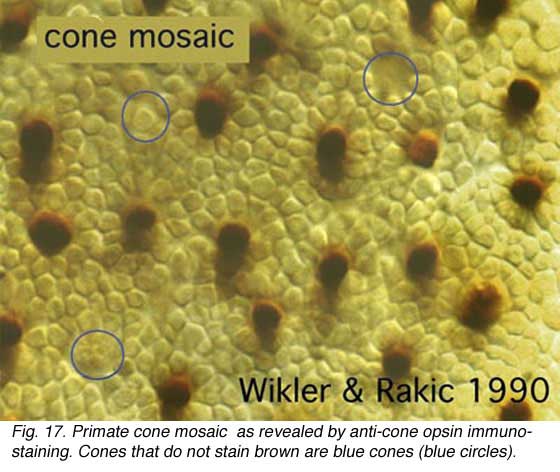 In Figure 17 in anti-visual pigment antibody stained tissue, the S-cones stand out as the cones that are not stained because the antibody recognizes only L- and M-cone visual pigment. i.e. the brown stained cone profiles are L- and M-cone types while the unstained profiles surrounded by blue circles are the S-cones (Wikler and Rakic, 1990).
In Figure 17 in anti-visual pigment antibody stained tissue, the S-cones stand out as the cones that are not stained because the antibody recognizes only L- and M-cone visual pigment. i.e. the brown stained cone profiles are L- and M-cone types while the unstained profiles surrounded by blue circles are the S-cones (Wikler and Rakic, 1990).
6. Morphology of the blue or S-cones
Recently, careful morphological studies have enabled us to distinguish the short wavelength specific (blue) cone from the medium and long wavelength specific cones in the human retina even without special antibody staining techniques (Ahnelt et al., 1987).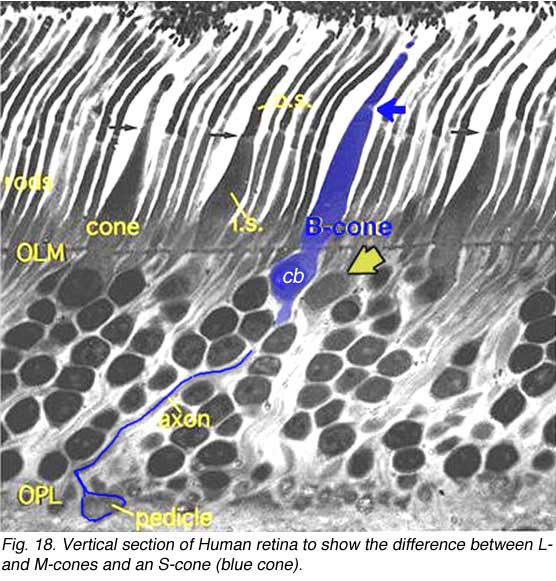 Thus, we now know that S-cones have longer inner segments that project further into subretinal space than longer wavelength cones (Fig 18). Their inner segment diameters do not vary much across the entire retina, thus they are fatter in the foveal area but thinner in the peripheral retina than longer wavelength cones. The S-cones also have smaller and morphologically different pedicles than the other two wavelength cones (Ahnelt et al., 1990). Furthermore, throughout the retina, the S-cones have a different distribution and do not fit into the regular hexagonal mosaic of cones typical of the other two types.
Thus, we now know that S-cones have longer inner segments that project further into subretinal space than longer wavelength cones (Fig 18). Their inner segment diameters do not vary much across the entire retina, thus they are fatter in the foveal area but thinner in the peripheral retina than longer wavelength cones. The S-cones also have smaller and morphologically different pedicles than the other two wavelength cones (Ahnelt et al., 1990). Furthermore, throughout the retina, the S-cones have a different distribution and do not fit into the regular hexagonal mosaic of cones typical of the other two types.
This is illustrated in the tangential section of the foveal cone mosaic (Fig. 19) where the hexagonal packing is distorted in many places by a larger-diameter cone (arrowed cones) breaking up the perfect mosaic into irregular subunits. The larger-diameter cones are S-cones. These cones have their lowest density in the foveal pit at 3-5% of the cones, reach a maximum density of 15% on the foveal slope (1 degree from the foveal pit) and then form an even 8% of the total population elsewhere in the retina (Ahnelt et al., 1987).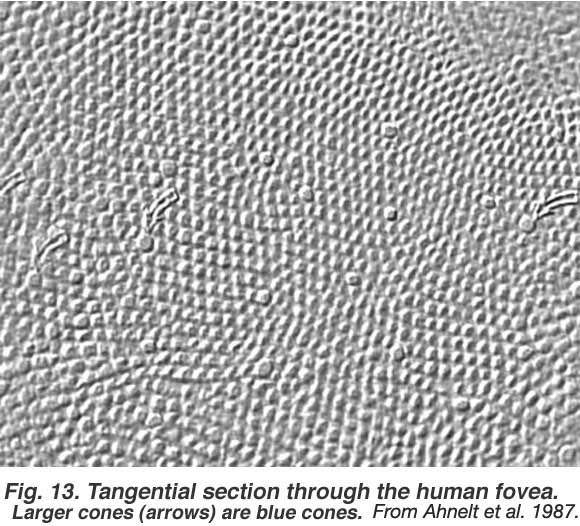 Analogous information concerning relative distributions of the M- and L-cones in the human retina are not easily available because we cannot tell them apart by morphological features or even by anti-visual pigment staining. In the monkey retina, Marc and Sperling (1977), performed a colored light-dependent histochemical staining technique on freshly excised monkey eyes. They found that L-cones (red) occur at about 33% of the cones throughout the retina, while M-cones (green) peak in the fovea at 64% and vary between 52% and 59% elsewhere in the retina. However, others have found the L-cones to outnumber the M-cones in fovea and perifoveal psychophysical testing paradigms (Cicerone and Nerger, 1989). The latest laser inferometry techniques (Roorda and Williams, 1999, Hofer et al. 2005), measuring the distribution of the red and green cones in the living human fovea, show there to be considerable variation amongst individuals. Some have an equal distribution of L- and M-cones, but others have a larger number of red cones even up to the ratio of 16 L-cones:1 M-cones. Both Roorda and Williams and Hofer et al’s (1999, 2005) data in human and Mollon and Bowmaker (1992) in monkey fovea show an irregular and patchwork nature of the L- and M-cone distribution (this is expanded on in the chapter on midget pathways, Figure 5).
Analogous information concerning relative distributions of the M- and L-cones in the human retina are not easily available because we cannot tell them apart by morphological features or even by anti-visual pigment staining. In the monkey retina, Marc and Sperling (1977), performed a colored light-dependent histochemical staining technique on freshly excised monkey eyes. They found that L-cones (red) occur at about 33% of the cones throughout the retina, while M-cones (green) peak in the fovea at 64% and vary between 52% and 59% elsewhere in the retina. However, others have found the L-cones to outnumber the M-cones in fovea and perifoveal psychophysical testing paradigms (Cicerone and Nerger, 1989). The latest laser inferometry techniques (Roorda and Williams, 1999, Hofer et al. 2005), measuring the distribution of the red and green cones in the living human fovea, show there to be considerable variation amongst individuals. Some have an equal distribution of L- and M-cones, but others have a larger number of red cones even up to the ratio of 16 L-cones:1 M-cones. Both Roorda and Williams and Hofer et al’s (1999, 2005) data in human and Mollon and Bowmaker (1992) in monkey fovea show an irregular and patchwork nature of the L- and M-cone distribution (this is expanded on in the chapter on midget pathways, Figure 5).
7. Densities of rods and cones in the human retina
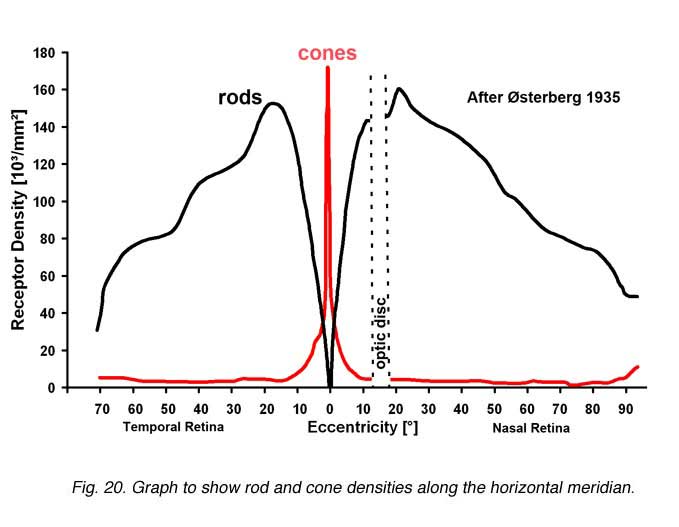
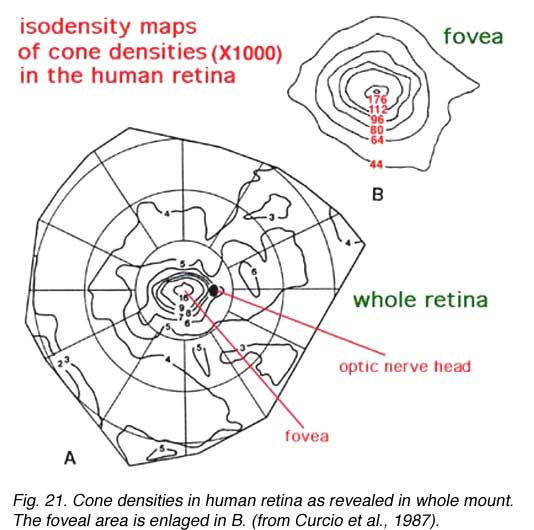
It is important for our understanding of the organization of the visual connections for us to know the spatial distribution of the different cell types in the retina. Photoreceptors, we know, are organized in a fairly exact mosaic. As we saw in the fovea, the mosaic is a hexagonal packing of cones. Outside the fovea, the rods break up the close hexagonal packing of the cones but still allow an organized architecture with cones rather evenly spaced surrounded by rings of rods. Thus in terms of densities of the different photoreceptor populations in the human retina, it is clear that the cone density is highest in the foveal pit and falls rapidly outside the fovea to a fairly even density into the peripheral retina (Figs. 20 and 21) (Osterberg, 1935; Curcio et al., 1987). There is a peak of the rod photoreceptors in a ring around the fovea at about 4.5 mm or 18 degrees from the foveal pit. The optic nerve (blind spot) is of course photoreceptor free (see below).
 |  |
{kind=link}
{kind=link}
8. Rods and Night Vision
Rods convey the ability to see at night, under conditions of very dim illumination. Animals with high densities of rods tend to be nocturnal, whereas those with mainly cones tend to be diurnal. The nature of dim light is important both to physicists and to biologists. In 1905 Einstein proposed that light propagated only in discrete irreducible packets or quanta (Einstein, 1905). This explained the non-classical features of the ‘photoelectric effect’, a process by which light releases electrons from metal surfaces, described by Heinrich Hertz in 1887 (Hertz, 1887). Rods are so sensitive that they actually detect single quanta of light, much as do the most sensitive of physical instruments. In 1942 Selig Hecht argued that human rods must be capable of detecting individual light quanta because light flashes so dim that only 1 in 100 rods were likely to absorb a quantum were yet reliably seen by careful observers (Hecht, 1942). A century after the original discovery of the photoelectric effect it has become possible to record directly the minute electrical voltages in rods induced by absorption of individual light quanta. An excellent example is shown in the suction electrode recordings of monkey rods by Schneeweis and Schnapf (1995) (Fig. 22). Each dot in the figure below represents delivery of a very dim pulse of light containing only a few quanta. Voltage responses appear to come in 3 sizes: none, small, and large, representing the detection of 0, 1 or 2 quanta in each flash. The granularity of response to dim light stimuli is evident.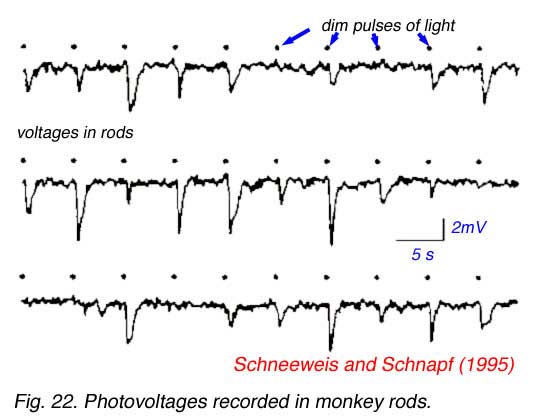 Rod sensitivity appears to be bought at a price, however, since rods are much slower to respond to light stimulation than cones. This is one reason why sporting events such as baseball become progressively more difficult as daylight fails. Both electrical recordings and human observations suggest that signals from rods may arrive as much as 1/10 second later than those from cones under lighting conditions where both can be simultaneously activated (MacLeod, 1972).
Rod sensitivity appears to be bought at a price, however, since rods are much slower to respond to light stimulation than cones. This is one reason why sporting events such as baseball become progressively more difficult as daylight fails. Both electrical recordings and human observations suggest that signals from rods may arrive as much as 1/10 second later than those from cones under lighting conditions where both can be simultaneously activated (MacLeod, 1972).
9. Ultrastructure of rod and cone synaptic endings
The job of the photoreceptor cell in the retina is to catch quanta of light in the visual pigment-containing membranes of the outer segment and pass a message, concerning numbers of quanta of light and sensitivities to the different wavelengths, to the next stage of integration and processing at the outer plexiform layer (see Transduction).
 |  |
{kind=link}
{kind=link}
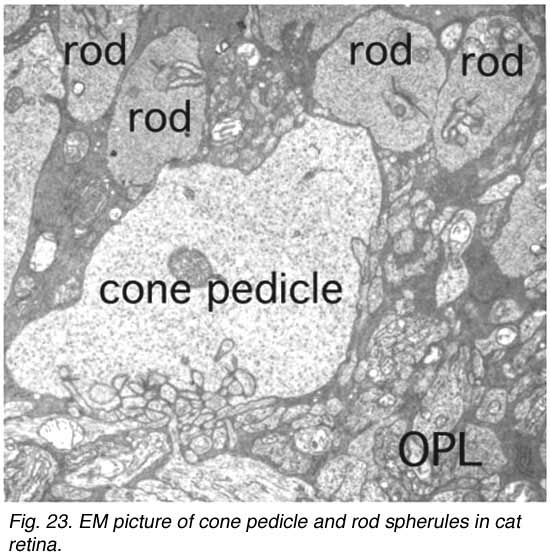
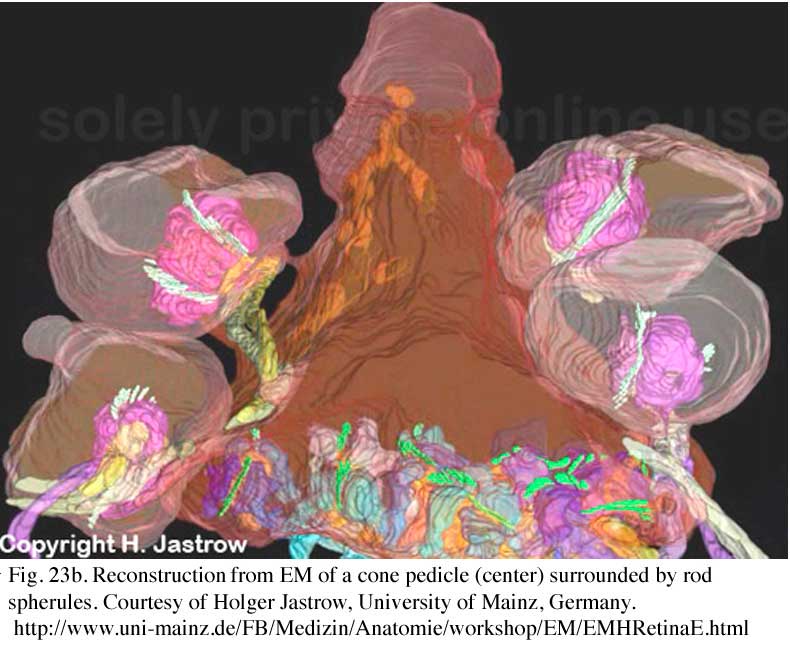
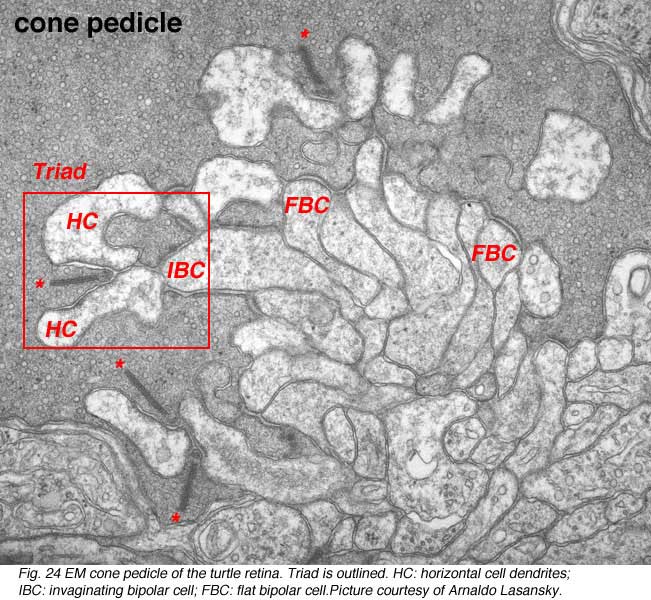
The information transmitting end of the cone cell is known as the pedicle and of the rod cell as the spherule. Cone pedicles are large, conical, flat end-feet (8-10 µm diameter) of the cone axon that lie more or less side by side on the same plane at the outer edge of the outer plexiform layer (OPL)(Figs. 23a and b). The more numerous rod spherules, in contrast, are small round enlargements of the axon (3-5 µm diameter) or even extensions of the cell body. They lie packed between and above the cone pedicles (Fig. 23a and b). Both photoreceptor types’ synaptic endings are filled with synaptic vesicles. At their synapses to second-order neurons (bipolar and horizontal cells), both rod spherules and cone pedicles exhibit dense structures known as synaptic ribbons pointing to the postsynaptic invaginated processes (asterisks in Fig. 24). In the cone pedicle approximately 30 of these ribbons occur and are associated with 30 “triads” of invaginated processes (Ahnelt et al., 1990). In the rod spherule 2 ribbons are associated with 4 invaginated second-order neurites while the cone pedicle delivers information to over a hundred second-order neurons (Fig. 23b).
 |  |
{kind=link}
{kind=link}
The cone “triad” of invaginated second-order processes consists typically of a central element which is a dendritic terminal of an invaginating bipolar cell (IBC), and two lateral elements which are dendritic terminals of horizontal cells (HC) (Figs. 24 and 27). In addition, other varieties of bipolar cell have dendrites making synaptic contacts on the under surface of the cone pedicle at what were first called flat contacts (FBC) (Missotten, 1965; Dowling and Boycott, 1966; Kolb, 1970) (Fig. 27), but then were better characterized and defined by Lasansky (1971) as basal junctions (Fig. 26).
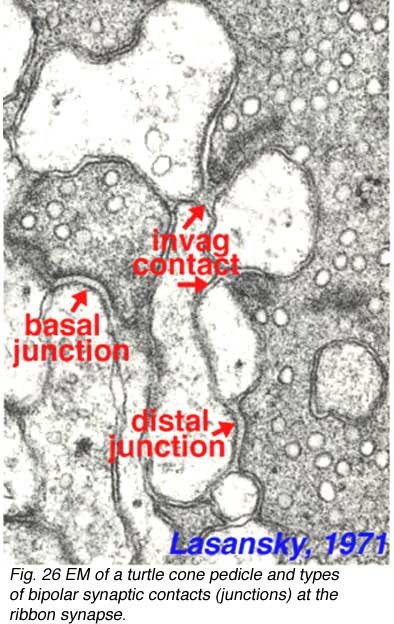 | 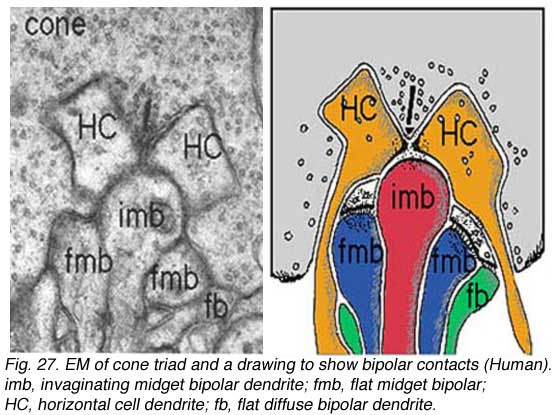 |
{kind=link}
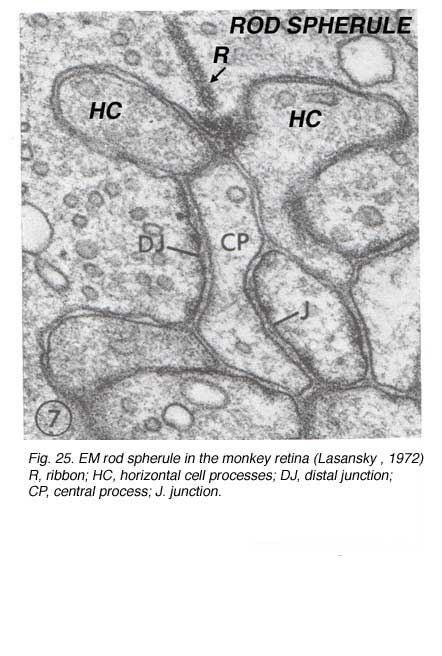
Rods spherules have only two synaptic ribbons associated with two lateral elements that are horizontal cell axon terminals (HC) and two central invaginating dendrites of rod bipolar cells (rb) (Missotten, 1965; Dowling and Boycott, 1966; Kolb, 1970). There are no basal junctions on rod spherules.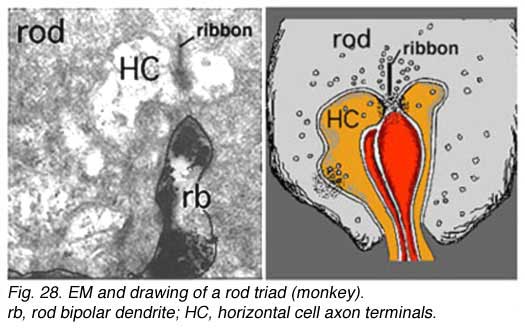
10. Interphotoreceptor contacts at gap junctions
There also appears to be a pathway for crosstalk between cones and cones and cones and rods in the human retina. Cone pedicles have small projections from their sides or bases that pass to neighboring rod spherules and cone pedicles. Where these projections, called telodendria, meet they have a specialized junction known to be typical of electrical synaptic transmission. These are minute gap junctions (Fig. 29) (Raviola and Gilula, 1975; Nelson et al., 1985).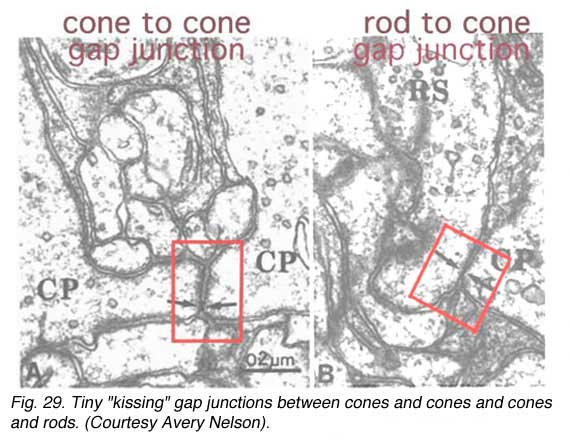 As many as 3-5 gap junctions occur on a single rod spherule from neighboring cone telodendria, and a single cone pedicle can have as many as 10 contacts to neighboring rods. Pedicles of S-cones do not have as many telodendrial gap junctions with either neighboring rods or cones (Ahnelt et al., 1990) and thus, this cone type remains relatively isolated in the cone mosaic, and, as we shall see later, remains isolated to the ganglion cell level too, due to connections with a specific ‘S-cone bipolar cell’.
As many as 3-5 gap junctions occur on a single rod spherule from neighboring cone telodendria, and a single cone pedicle can have as many as 10 contacts to neighboring rods. Pedicles of S-cones do not have as many telodendrial gap junctions with either neighboring rods or cones (Ahnelt et al., 1990) and thus, this cone type remains relatively isolated in the cone mosaic, and, as we shall see later, remains isolated to the ganglion cell level too, due to connections with a specific ‘S-cone bipolar cell’.
Direct interactions between different functional classes of photoreceptors were not anticipated based on any known or theoretical needs of the visual system. In fact such connections would appear to degrade spatial resolution, and, potentially, color perception by mixing signals from photoreceptors at different locations or with different photopigments. Nonetheless, in agreement with anatomical findings, mammalian cones appear to carry rod signals. The slow wave forms of rod signals are easily distinguished from the rapid wave forms of cone signals in voltage recordings from single cones. Examples of voltage responses from monkey cones are illustrated in Figure 30 (Schneeweis and Schnapf, 1995). Voltage records resulting from balanced red and green stimuli are illustrated.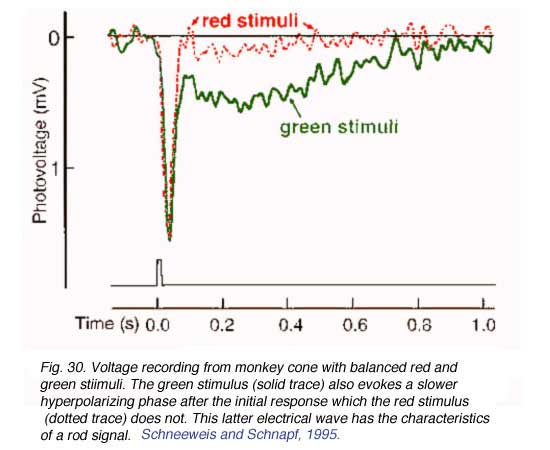 Both records exhibit the same initial peak hyperpolarization in response to the brief stimulus, however the green stimulus (solid trace) also evokes a slower hyperpolarizing phase after the initial response which the red stimulus (dotted trace) does not (Fig. 30). This latter electrical wave has the characteristics of a rod signal. Such signals have also been observed in cat cones (Nelson, 1977). One theory of the utility of this arrangement is that it allows rods to utilize neural pathways devoted to both cones and rods in sending visual information to the inner plexiform layer. Cone pathways may be tuned to faster temporal characteristics than rod pathways, and so by utilizing both pathways, rods may transmit a wider bandwidth of temporal information. There is evidence for two rod pathways with different dynamic signatures in perceptual experiments (Sharpe et al, 1989). Although the functional role of interreceptor junctions is still a matter of debate, they perhaps serve as a philosophical warning to studies of biological sensory systems: Not even the receptor cells themselves stand in isolation of the activity and influence of neighboring neurons.
Both records exhibit the same initial peak hyperpolarization in response to the brief stimulus, however the green stimulus (solid trace) also evokes a slower hyperpolarizing phase after the initial response which the red stimulus (dotted trace) does not (Fig. 30). This latter electrical wave has the characteristics of a rod signal. Such signals have also been observed in cat cones (Nelson, 1977). One theory of the utility of this arrangement is that it allows rods to utilize neural pathways devoted to both cones and rods in sending visual information to the inner plexiform layer. Cone pathways may be tuned to faster temporal characteristics than rod pathways, and so by utilizing both pathways, rods may transmit a wider bandwidth of temporal information. There is evidence for two rod pathways with different dynamic signatures in perceptual experiments (Sharpe et al, 1989). Although the functional role of interreceptor junctions is still a matter of debate, they perhaps serve as a philosophical warning to studies of biological sensory systems: Not even the receptor cells themselves stand in isolation of the activity and influence of neighboring neurons.
11. References
- Adler AJ, Martin KJ. Retinol-binding in bovine interphotoreceptor matrix. Biochem Biophys Res Commun. 1982;108:1601–1608. [PubMed]
- Ahnelt PK, Kolb H, Pflug R. Identification of a subtype of cone photoreceptor, likely to be blue sensitive, in the human retina. J Comp Neurol. 1987;255:18–34. [PubMed]
- Ahnelt PK, Keri C, Kolb H. Identification of pedicles of putative blue sensitive cones in human and primate retina. J Comp Neurol. 1990;293:39–53.[PubMed]
- Ammermuller, J. and H. Kolb (1996) Functional architecture of the turtle inner retina. Prog. Ret. & Eye Res. 15 (2), 393-433.
- Anderson DH, Fisher SK. The photoreceptors of diurnal squirrels: outer segment structure, disc shedding, and protein renewal. J Ultrastruct Res.1976;55:119–141. [PubMed]
- Archer S. Molecular biology of visual pigments. In: Djamgoz MBA, Archer SN, Vallerga S, editors. Neurobiology and clinical aspects of the outer retina. London: Chapman & Hall; 1995. p. 79-104.
- Baylor DA, Nunn BJ, Schnapf JL (1987) Spectral sensitivity of cones of the monkey Macaca fascicularis. J Physiol. 390:145-60. [PubMed]
- Besharse JC. The daily light-dark cycle and rythmic metabolism in the photoreceptor-pigment epithelial complex. Prog Ret Res. 1982;1:81–124.
- Bowmaker JK, Dartnall HJ (1980) Visual pigments of rods and cones in a human retina. J Physiol. 298: 501-11. [PubMed]
- Bowmaker JK, Dartnall HJ, Mollon JD (1980) Microspectrophotometric demonstration of four classes of photoreceptor in an old world primate, Macaca fascicularis. J Physiol. 298: 131-43. [PubMed]
- Chader GJ. Interphotoreceptor retinoid-binding protein (IRBP): a model protein for molecular biological and clinically relevant studies. Invest Ophthal Vis Sci.1989;30:7–22. [PubMed]
- Cicerone CM, Nerger JL. The relative numbers of long-wavelength-sensitive to middle-wavelength-sensitive cones in the human fovea centralis. Vision Res.1989;29:115–128. [PubMed]
- Curcio CA, Sloan KR, Packer O, Hendrickson AE, Kalina RE. Distribution of cones in human and monkey retina: individual variability and radial asymmetry.Science. 1987;236:579–582. [PubMed]
- DeMonasterio FM, Schein SJ, McCrane EP. Staining of blue sensitive cones of the Macaque retina by fluorescent dye. Science. 1981;213:1278–1281.[PubMed]
- Deretic D, Papermaster DS. The role of small G-proteins in the transport of newly synthesized rhodopsin. Prog Ret Eye Res. 1995;14:249–265.
- Dobelle WH, Marks WB, MacNichol EF Jr. (1969) Visual pigment density in single primate foveal cones. Science. 166(3912): 1508-10. [PubMed]
- Dowling JE, Boycott BB. Organization of the primate retina: electron microscopy. Proc R Soc B Lond. 1966;166:80–111.
- Einstein, A., On a Heuristic Viewpoint Concerning the Production and Transformation of Light. Annalen der Physik 1905. 17(6): p. 132-148.
- Gouras P. Color vision. Prog Ret Res. 1984;3:227–261.
- Hargrave PA, McDowell JH, Feldmann RJ, Atkinson PH, Rao JKM, Argos P. Rhodopsin’s protein and carbohydrate structure: selected aspects. Vision Res.1984;24:1487–1499. [PubMed]
- Hargrave PA, McDowell JH. Rhodopsin and phototransduction. Int Rev Cytol. 1992;137B:49–97. [PubMed]
- Hecht S, Schlaer S, Pirenne MH. Energy, quanta and vision. J Gen Physiol. 1942;25:819–840. [PubMed]
- Hertz, H., Ueber den Einfluss des ultravioletten Lichtes auf die electrische Entladung. Annalen der Physik, 1887. 267(8): p. 983-1000.
- Hofer H, Carroll J, Neitz J, Neitz M, Williams DR. Organization of the human trichromatic cone mosaic. J Neurosci. 2005;25:9669–9679. [PubMed]
- Jastrow H. Electron microscopic atlas of cells, tissues and organs in the internet. Available from: http://www.uni-mainz.de/FB/Medizin/Anatomie/workshop/EM/EMHRetinaE.html.
- Kawamura S. Photransduction, excitation and adaptation. In: Djamgoz MBA, Archer SN, Vallerga S, editors. Neurobiology and clinical aspects of the outer retina. London: Chapman & Hall; 1995. p. 105-131.
- Kolb H. Organization of the outer plexiform layer of the primate retina: electron microscopy of Golgi-impregnated cells. Phil Trans R Soc B Lond.1970;258:261–283.
- Kolb H, Lipetz LE. The anatomical basis for colour vision in the vertebrate retina. In: Gouras P, editor. Vision and visual dysfunction. Vol. 6, The perception of colour. London: Macmillan Press Ltd.; 1991. p. 128-145.
- Lasansky A. Synaptic organization of cone cells in the turtle retina. Phil Trans R Soc B. 1971;262:365–381.
- LaVail MM. Rod outer segment disc shedding in rat retina: relationship to cyclic lighting. Science. 1976;194:1071–1074. [PubMed]
- MacLeod DIA. Rods cancel cones in flicker. Nature. 1972;235:173–174. [PubMed]
- Marc RE, Sperling HG. Chromatic organization of primate cones. Science. 1977;196:454–456. [PubMed]
- Missotten L. The ultrastructure of the human retina. Brussels: Arscia Uitgaven N.V.; 1965.
- Mollon JD, Bowmaker JK. The spatial arrangement of cones in the primate fovea. Nature. 1992;360:677–679. [PubMed]
- Nathans J, Piantanida TP, Eddy RL, Shows TB, Hogness DS. (1986) Molecular genetics of inherited variation in human color vision. Science. 232, 203-10. [PubMed]
- Nelson R. Cat cones have rod input: a comparison of the response properties of cones and horizontal cell bodies in the retina of the cat. J Comp Neurol.1977;172:109–136. [PubMed]
- Nelson R, Lynn T, Dickinson-Nelson A, Kolb, H. Spectral mechanisms in cat horizontal cells. In: Gallego A, Gouras P, editors. Neurocircuitry of the retina: a Cajal memorial. 1985; p. 109-121.
- Normann RA, Perlman I, Hallet PE. Cone photoreceptor physiology and cone contributions to colour vision. In: Gouras P, editor. Vision and visual dysfunction. Vol. 6, The perception of colour. London: Macmillan Press Ltd.; 1991. p. 146-162.
- Osterberg G. Topography of the layer of rods and cones in the human retina. Acta Ophthal Suppl. 1935;6:1–103.
- Papermaster DS, Schneider BG, Besharse JC. Vesicular transport of newly synthesized opsin from the Golgi apparatus toward the rod outer segment. Invest Ophthal Vis Sci. 1985;26:1386–1404. [PubMed]
- Raviola E, Gilula NB. Intramembrane organization of specialized contacts in the outer plexiform layer of the retina: a freeze-fracture study in monkeys and rabbits. J Cell Biol. 1975;65:192–222. [PubMed]
- Roorda A, Williams DR. The arrangement of the three cone classes in the living human eye. Nature. 1999;397:520–522. [PubMed]
- Schneeweis DM, Schnapf JL. Photovoltage of rods and cones in the macaque retina. Science. 1995;268:1053–1055. [PubMed]
- Sharpe LT, Stockman A, MacLeod DIA. Rod flicker perception: scotopic duality, phase lags and destructive interference. Vision Res. 1989;29:1539–1559.[PubMed]
- Steinberg RH, Wood I, Hogan MJ. Pigment epithelial ensheathment and phagocytosis of extra foveal cones in the human retina. Phil Trans R Soc B.1977;277:459–474. [PubMed]
- Steinberg RH, Fisher SK, Anderson DH. Disc morphogenesis in vertebrate photoreceptors. J Comp Neurol. 1980;190:501–518. [PubMed]
- Stryer L. Visual excitation and recovery. J Biol Chem. 1991;266:10711–24. [PubMed]
- Szel A, Diamanstein T, Rohlich P. Identification of blue-sensitive cones in the mammalian retina by antivisual pigment antibody. J Comp Neurol.1988;273:593–602. [PubMed]
- Wikler KC, Rakic P. Distribution of photoreceptor subtypes in the retina of diurnal and nocturnal primates. J Neurosci. 1990;10:3390–3401. [PubMed]
- Yau K-W. Phototransduction mechanisms in retinal rods and cones. Invest Ophthal Vis Sci. 1994;35:9–32. [PubMed]
- Young RW. The renewal of rod and cone outer segments in the rhesus monkey. J Cell Biol. 1971;49:303–318. [PubMed]
- Young RW. Visual cells and the concept of renewal. Invest Ophthalmol. 1976;15:700–725.