
When looking into someone’s eyes, we can easily see several structures:
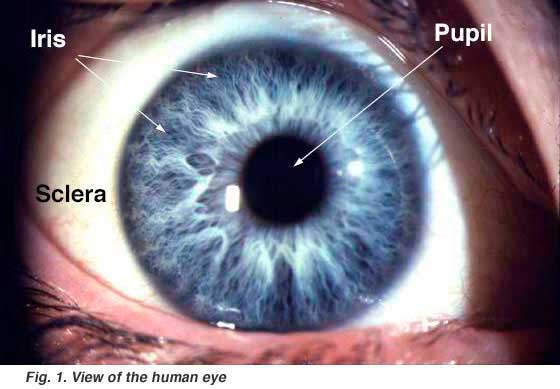
- A black-looking aperture, the pupil, that allows light to enter the eye (it appears dark because of the absorbing pigments in the retina) (Fig. 1).
- A colored circular muscle, the iris, which is beautifully pigmented giving us our eye’s color (the central aperture of the iris is the pupil) (Fig. 1). This circular muscle controls the size of the pupil so that more or less light, depending on conditions, is allowed to enter the eye. Eye color, or more correctly, iris color is due to variable amounts of eumelanin (brown/black melanins) and pheomelanin (red/yellow melanins) produced by melanocytes. More of the former is in brown eyed people and of the latter in blue and green-eyed people. The Melanocortin-1 Receptor Gene is a regulator of eumelanin production and is located on chromosome(MCIR) 16q24.3. Point mutations in the MCIR gene will affect melanogenesis. The presence of point mutations in the MCIR gene alleles is a common feature in light skinned and blue/green eyed people (J.A.W. Metzelaar-Blok et al. 2001; P. Valverde et al. 1995).
- A transparent external surface, the cornea, that covers both the pupil and the iris. This is the first and most powerful lens of the optical system of the eye and allows, together with the crystalline lens the production of a sharp image at the retinal photoreceptor level.
- The “white of the eye”, the sclera,which forms part of the supporting wall of the eyeball. The sclera is continuous with the cornea. Furthermore this external covering of the eye is in continuity with the dura of the central nervous system.
When we remove the eye from the orbit, we can see that the eye is a slightly asymmetrical sphere with an approximate sagittal diameter or length of 24 to 25 mm. and a transverse diameter of 24 mm. It has a volume of about 6.5 cc
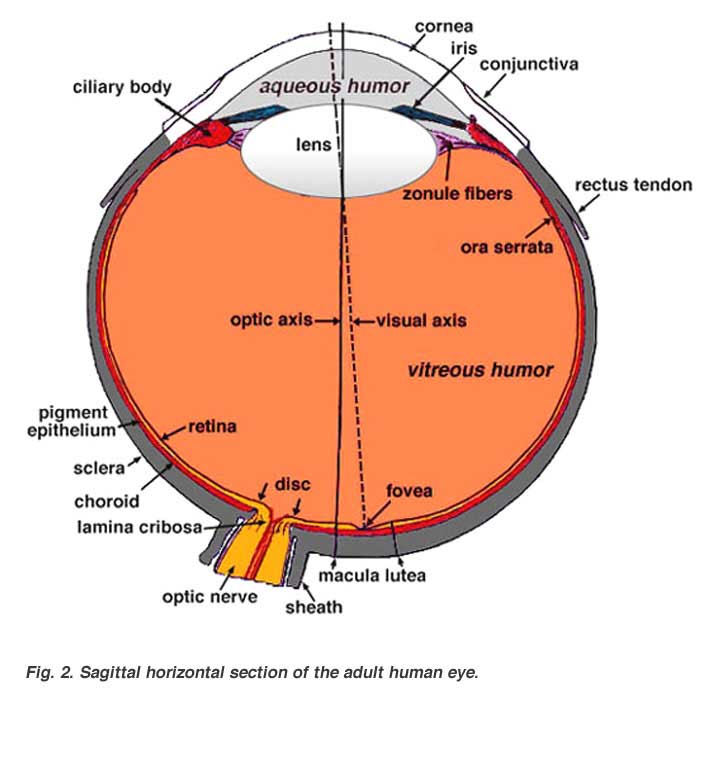
A cross-sectional view of the eye (Fig. 2) shows:
- Three different layers
- The external layer, formed by the sclera and cornea
- The intermediate layer, divided into two parts: anterior (iris and ciliary body) and posterior (choroid)
- The internal layer, or the sensory part of the eye, the retina
- Three chambers of fluid: Anterior chamber (between cornea and iris), Posterior chamber (between iris, zonule fibers and lens) and the Vitreous chamber (between the lens and the retina). The first two chambers are filled with aqueous humor whereas the vitreous chamber is filled with a more viscous fluid, the vitreous humor.
- The sagittal section of the eye also reveals the lens which is a transparent body located behind the iris. The lens is suspended by ligaments (called zonule fibers) attached to the anterior portion of the ciliary body. The contraction or relaxation of these ligaments as a consequence of ciliary muscle actions, changes the shape of the lens, a process called accommodation that allows us to form a sharp image on the retina.
Light rays are focussed through the transparent cornea and lens upon the retina. The central point for image focus (the visual axis) in the human retina is the fovea. Here a maximally focussed image initiates resolution of the finest detail and direct transmission of that detail to the brain for the higher operations needed for perception. Slightly more nasally than the visual axis is the optic axis projecting closer to the optic nerve head. The optic axis is the longest sagittal distance between the front or vertex of the corna and the furthest posterior part of the eyeball. It is about the optic axis that the eye is rotated by the eye muscles. Some vertebrate retinas have instead of a fovea, another specialization of the central retina, known as an area centralis or a visual streak.
1.Extraocular muscles
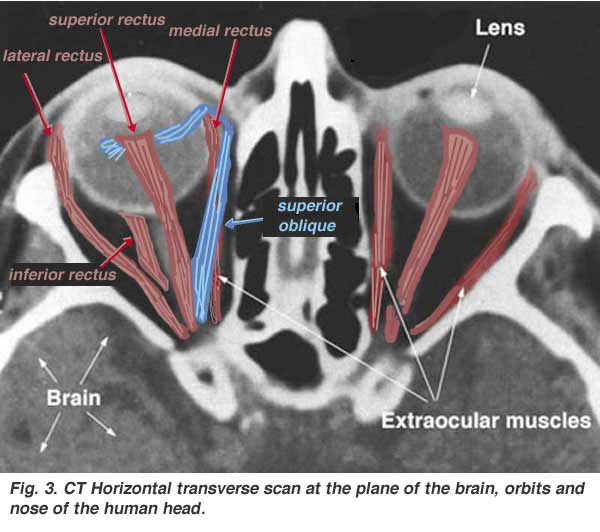
Each eyeball is held in position in the orbital cavity by various ligaments, muscles and fascial expansions that surround it (see Fig. 3).
Inserted into the sclera are three pairs of muscles (6 muscles altogether). Two pairs are rectus muscles running straight to the bony orbit of the skull orthogonal to each other (the superior rectus, the inferior rectus, the lateral rectus and the medial rectus muscles). A further pair of muscles, the oblique muscles (superior oblique and inferior oblique) are angled as the name implies obliquely. These muscles, named extraocular muscles rotate the eyeball in the orbits and allow the image to be focussed at all times on the fovea of central retina.
2.Development of the eye
The retina is a part of the central nervous system and an ideal region of the vertebrate brain to study, because like other regions of the central nervous system, it derives from the neural tube. The retina is formed during development of the embryo from optic vesicles outpouching from two sides of the developing neural tube. The primordial optic vesicles fold back in upon themselves to form the optic cup with the inside of the cup becoming the retina and the outside remaining a single monolayer of epithelium known as the retinal pigment epithelium (Fig, 4, 3). Initially both walls of the optic cup are one cell thick, but the cells of the inner wall divide to form a neuroepithelial layer many cells thick: the retina


Further changes in retinal morphology are accomplished by simultaneous formation of multiple complex intercellular connections. Thus by 5 months of gestation (Fig. 5) most of the basic neural connections of the retina have been established (Mann, 1964).
The functional synapses are made almost exclusively in the two plexiform layers and the perikarya of the nerve cells are distributed in the three nuclear layers.
Photoreceptor cell maturation begins with the formation of outer segments (OS) containing visual pigment from multiple infoldings of the plasma membrane of each cell. Outer segment formation proceeds, and the eye becomes sensitive to light at about 7 months’ gestation.
The final portion of the sensory retina to mature is the fovea, where the ganglion cell layer thickening begins during midgestation. The outer nuclear layer is also wider here than elsewhere in the retina and consists almost entirely of developing cone cells. The ganglion cell nuclei migrate radially outwards in a circle, leaving the fovea free of ganglion cell nuclei. Cell-cell attachments persist, however and foveal cone cells alter their shape to accomodate the movement of ganglion cells. Foveal development continues with cell rearrangements and alteration in cone shape until about 4 years after birth (Hendrickson and Yondelis, 1984; Curcio and Hendrickson, 1991).
Surface membranes cover the eye cup and develop into lens, iris and cornea with the three chambers of fluid filled with aqueous and vitreous humors (Fig. 6).
3.Summary
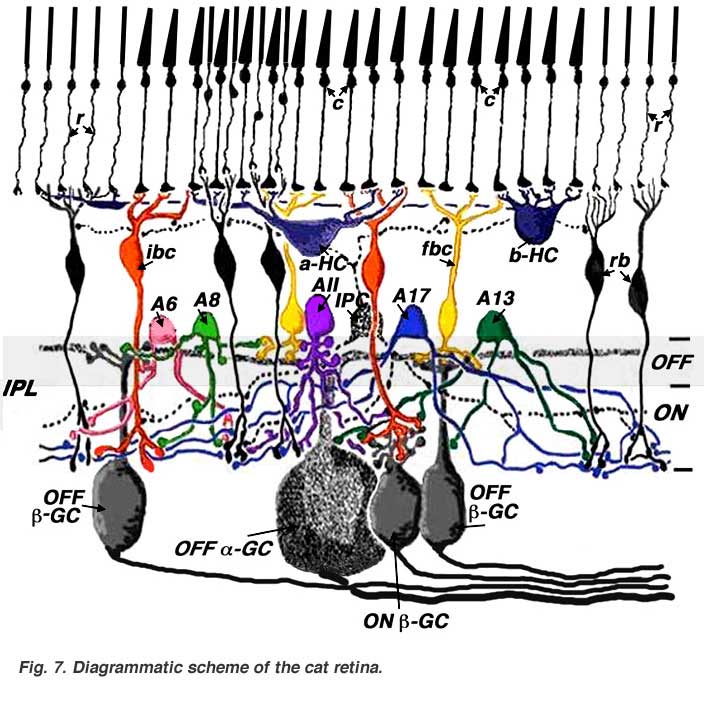
In following chapters, we will describe in greater detail the individual nerve cells that make up the retina and the functional pathways into which these neurons are organized. Eventually, we will progress to a stage where we can appreciate the summary diagrams below (Figs. 7 and 8) that show the functional wiring of two well-understood mammalian retinas, namely cat and primate retinas.
 |  |
4.References
- Curcio CA, Hendrickson AE. Organization and development of the primate photoreceptor mosaic. Prog Ret Ret. 1991;10:89–120.
- Hendrickson AE, Youdelis C. The morphological development of the human fovea. Ophthalmology. 1984;91:603–612. [PubMed]
- Mann I. The development of the human eye. New York: Grune and Stralton; 1964.
- Metzelaar-Blok JAW, ter Huurne JA, Hurks HM, Keunen JE, Jager MJ, Gruis NA. (2001) Characterization of melanocortin-1 receptor gene variants in uveal melanoma patients. Invest Ophthalmol Vis Sci. 42:1951-1954. [PubMed]
- Ogden TE. Retina: basic science and inherited retinal disease. Vol. 1. St. Louis: The CV Mosby Co.; 1989.
- Valverde P, Healy E, Jackson I, Rees JL, Thody AJ. (1995) Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat Genet.;11:328-330. [PubMed]